皮肤微生态是指由细菌、真菌、病毒等微生物与皮肤表面的组织、细胞、分泌物以及微环境共同组成的微生态系统,能通过调控皮肤屏障功能和调节免疫来维持皮肤的健康状态。近年来,皮肤微生态的稳态维系机制受到广泛关注。研究发现,钙离子通道作为细胞膜信号传导的核心元件,通过调控表皮分化、促进角质层脂质有序排列及加强紧密连接蛋白的表达,在维持皮肤物理屏障完整性中发挥关键作用。这种屏障功能的稳定性又与微生物群的定植抗性相关,完整的皮肤屏障可防止病原体入侵,同时通过调节pH、分泌抗菌肽等机制为共生菌提供适宜的生态位。
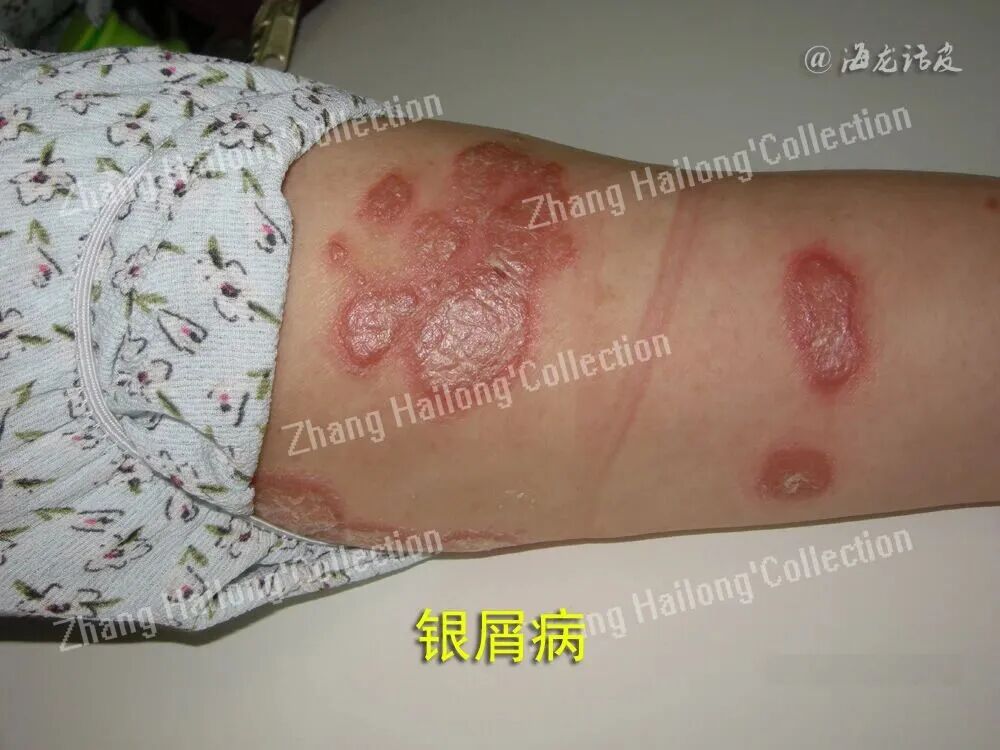
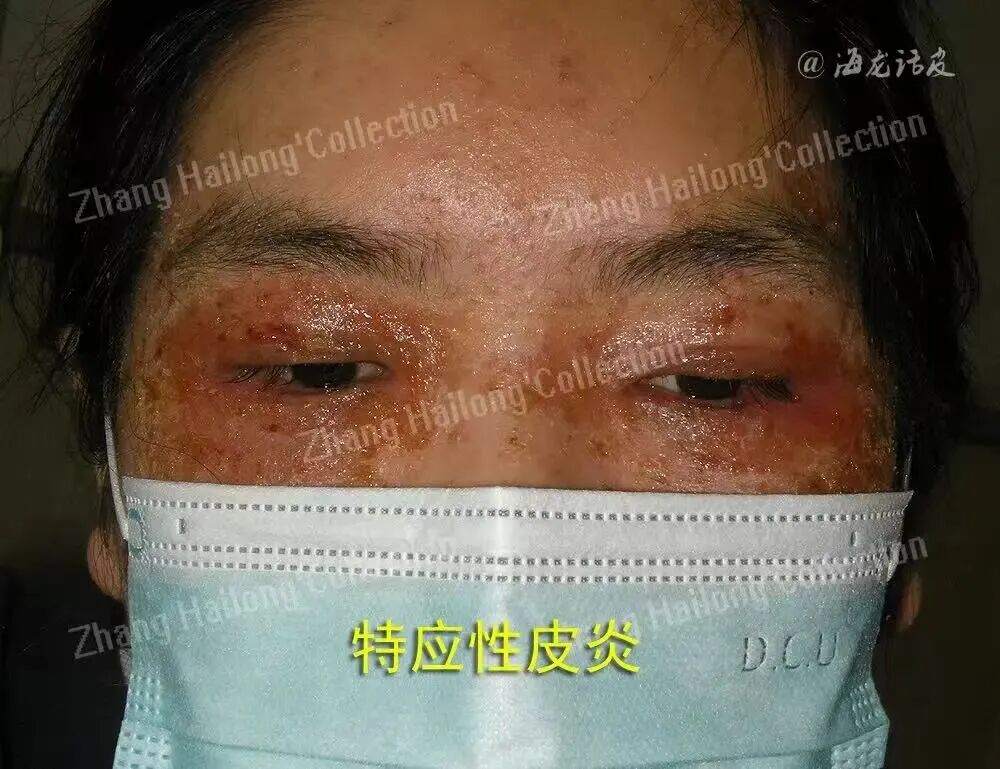
银屑病、特应性皮炎、痤疮和慢性伤口等多种炎症性和感染性疾病的发生与皮肤微生态失衡密切相关。银屑病皮损区表皮钙离子梯度紊乱导致角质形成细胞过度增殖,同时金黄色葡萄球菌(Staphylococcus aureus)定植的增加与TRPV3通道介导的白介素(interleukin,IL)-23/辅助性T细胞(helper T cell,Th)17炎症轴激活显著相关;特应性皮炎患者皮肤屏障缺陷部位因机械敏感通道的电通道组分1(piezo-type mechanosensitive ion channel component 1,Piezo1)过度激活,引发IL-31过量分泌,进而破坏表皮葡萄球菌(Staphylococcus epidermidis)的共生平衡。这些现象提示,钙离子通道可能通过“屏障−免疫−微生物”三位一体的调控网络来影响皮肤微生态。皮肤微生态失衡主要分为内源性和外源性两种类型。内源性失衡表现为固有菌群比例失调、移位定植和机会性感染;外源性失衡则由致病菌入侵引发菌群竞争性排斥,导致常驻菌群数量锐减。此外,紫外线和化妆品中的抗菌剂等也可能破坏皮肤微生态平衡。
目前,在化妆品行业中,人们在微生态护肤品中添加益生元,旨在通过调节皮肤微生态的平衡来改善皮肤状态,从而减少皮肤疾病的发生。添加果胶能够抑制常见皮肤病原体金黄色葡萄球菌的生长;而添加丁酸钠可以调节角质细胞的凋亡来预防银屑病。然而,关于影响和改变皮肤微生态微观因素的研究仍然较少。众所周知,钙离子通道蛋白作为细胞膜上的主要蛋白,参与多种生理过程,包括细胞增殖、分化、凋亡以及免疫调节等。关于钙离子通道调控皮肤微生态的研究近年来大幅增长,但是对于钙离子通道在其中的作用机制仍缺乏系统的归纳和阐述。
皮肤微生态概述
1.1、组成与特点:
微生物覆盖人类皮肤表面的面积约2m2。细菌、病毒和真菌定植于皮肤表面的内陷和褶皱区域,这些微生物在维持皮肤组织的稳态和局部免疫方面发挥着重要作用。研究表明,多种因素共同调控皮肤表面的微生态系统,主要包括皮肤的温度、湿度、pH值、抗菌肽的分泌以及皮肤脂质的组成和分布等。(1)皮肤表面温度的波动能够激活瞬时受体电位香草酸亚型1(transient receptor potential vanillic acid subtype1,TRPV1)通道,从而诱导角质形成细胞释放抗菌肽-β-防御素2。如在银屑病皮损区域,TRPV1的过度激活会导致局部温度升高超过2℃,这使得表皮葡萄球菌的相对丰度降低50%,而金黄色葡萄球菌显著增加。(2)皮肤pH值的稳态与VGCC的Cav1.3密切相关。当pH值超过6.0时,Cav1.3通道的开放概率增加,触发ATP12A质子泵的活化以恢复酸性微环境,从而抑制马拉色菌属(Malassezia)的β-葡萄糖苷酶的活性和脂质分解的能力。(3)皮肤的湿度平衡主要通过MS的Piezo1调控紧密连接蛋白的表达,得以维持表皮的水合状态,敲除Piezo1蛋白会导致经皮失水率上升,并增强痤疮丙酸杆菌(Propionibacterium acnes)生物膜的形成能力。此外,汗液中的D-丝氨酸作为离子型谷氨酸受体NMDAR的共激动剂,通过Cav3.2通道调节朗格汉斯细胞IL-1β的分泌,维持表皮葡萄球菌与耐甲氧西林金黄色葡萄球菌(methicillin-resistant Staphylococcus aureus)之间的动态平衡。
另外,皮肤附属器形成差异化微环境。毛囊、汗腺和皮脂腺等结构各自承载着独特的菌群,其菌群组成和功能特征呈现出显著的空间异质性。毛囊-皮脂腺单位形成的低氧微环境,为痤疮丙酸杆菌等兼性厌氧菌提供了优势生态位。富含脂质的皮脂不仅为微生物提供了丰富的营养来源,还促进了诸如痤疮丙酸杆菌和马拉色菌属等微生物在皮肤表面的扩张。皮肤表面微生物的扩张受到多种环境因素的共同调控,包括宿主皮肤表面角质细胞的持续脱落和汗液的分泌,这些过程共同作用于皮肤微环境,使得微生物群的扩张趋向动态平衡。
1.2、功能与重要性:
皮肤微生物群包含上千种细菌,主要包括放线菌门(Actinobacteria)、厚壁菌门(Firmicutes)和变形菌门(Proteobacteria)等,而病毒、真核生物及其他生物所占比例较小。这些微生物与皮肤健康密切相关。分析皮肤微生物组发现,银屑病、特应性皮炎和痤疮等皮肤疾病的发生源于微生物群内部菌种间精细平衡的失衡。这种失衡可能表现为某些有益微生物的减少或缺失,同时伴随着潜在有害微生物的过度繁殖,从而破坏了皮肤表面的微生态平衡。皮肤微生物群通过与皮肤细胞的直接或间接作用,参与皮肤细胞的分化和再生,帮助维持皮肤屏障功能。它们能够刺激皮肤和免疫细胞,提高对病原微生物的免疫反应。这些作用包括产生抗菌肽、调节免疫以及升高紧密连接蛋白的表达等,这些效应部分源于角质形成细胞和其他局部免疫细胞先天免疫受体的激活。此外,一些潜在的抗菌分子,如天然抗生素和共生微生物分泌的游离脂肪酸等,也有助于保护皮肤屏障。
1.3、平衡的机制:
皮肤微生态稳态是维持皮肤屏障功能的核心要素,但其稳态调控网络仍未完全解析。这主要是因为皮肤微生物组的构成具有高度的个体差异性,其复杂性不仅体现在微生物与宿主皮肤环境之间的相互作用,还涉及微生物种群内部以及不同种群之间的竞争关系。这些因素相互交织,使得整个系统尤为复杂。以往的研究聚焦于特定微生物对皮肤屏障功能的潜在破坏作用。以金黄色葡萄球菌为例,其通过生物膜的形成和抗原变异等策略逃避免疫清除,实现在皮肤的持续定植。此外,该菌还能分泌毒素,这些毒素具有扰乱适应性免疫应答的功能,进一步削弱皮肤屏障的完整性,并激发病理性炎症反应。
更为复杂的是,金黄色葡萄球菌与表皮葡萄球菌之间存在协同作用,它们通过酚溶性调节素α(phenol-soluble modulin α)这一信号分子,共同促进过度的蛋白水解酶活性,并增加金黄色葡萄球菌staphopainA和staphopainB蛋白酶的表达。在丝氨酸蛋白酶抑制剂缺失时,这些微生物的协同作用显著加剧皮肤炎症并破坏皮肤屏障。但是,宿主皮肤在面对外源性病原体侵袭时展现出了强大的防御机制,凸显了皮肤屏障功能的重要性与其作为人体的第一道防线的重要地位,这些发现为理解宿主防御机制奠定了基础。研究表明,吞噬细胞和嗜中性粒细胞在清除病原体方面发挥关键作用,为抗感染治疗提供了理论依据。此外体液免疫与细胞免疫在病原体清除中的协同作用,为疫苗研发和免疫治疗提供了新的思路。这些发现极大地丰富了我们对皮肤免疫防御机制的理解。
参与皮肤微生态调节的钙信号
钙离子作为细胞功能的通用信号,在宿主与微生物相互作用的过程中发挥着重要作用。研究表明,细菌感染或脂多糖(lipopolysaccharide,LPS)通过激活MS与Toll样受体(Toll-like receptor,TLR)4,重塑F−肌动蛋白的结构并增强吞噬功能。机制上,LPS通过TLR4激活MS介导的钙内流,驱动钙调蛋白依赖性蛋白激酶II(Calmodulin-dependent protein kinase II,CaMK II)-哺乳动物STE20样激酶1/2(mammalian STE20-like kinase1/2,Mst1/2)-Rac信号轴的激活,促进病原体的吞噬和清除。抑制CaMKII,或敲除Mst1/2或Rac家族小GTPase1(Rac family small GTPase 1,Rac1)导致巨噬细胞杀菌活性降低,且该通路与P2X7受体介导的ATP-Ca2+信号存在正向协同关系,即LPS触发的初期钙内流促进ATP的释放,后者通过激活嘌呤能受体进一步增强钙振荡幅度,形成自我强化回路。
这种时空耦合的钙信号特征能够特异性激活活化T细胞核因子的转录程序,诱导抗菌肽hBD-2的表达,同时通过Rac依赖性膜皱褶的形成来增强吞噬效率。宿主细胞通过感知细菌感染引发的钙离子信号及其下游传导通路的变化,形成先天性免疫应答机制,但在特定病理条件下,这一机制可能为病原体感染提供可乘之机。例如,在细胞侵袭过程中,与宿主细胞的接触会激活侵袭素IpaB和IpaC,这些因子插入宿主细胞质膜并触发导致肌动蛋白聚合的初始信号,同时允许其他细菌效应因子进入胞质,进而通过细胞骨架重塑,触发细胞内钙离子浓度的瞬时峰值。此外,肌动蛋白与磷脂酶C的协同作用,打开连接蛋白半通道,促使ATP释放并增强钙信号,进一步加速细菌的侵袭扩散。钙离子稳态的破坏可能普遍加剧病原菌的侵袭能力。皮肤表面的共生菌虽然通常不会引发炎症,但是一旦其侵入真皮层则会触发炎症反应。
钙离子通过抑制金黄色葡萄球菌的定植与增殖,增强皮肤对该病原体的抵御能力。该信号通过激活钙依赖性半胱氨酸蛋白酶,降解E-钙黏蛋白,破坏上皮屏障的完整性。值得注意的是,病原菌诱导的PLCγ-IP3通路会与宿主的防御性CaMKII通路形成动态拮抗, 即当胞内钙浓度上升时,钙调蛋白与CaMKII的解离效率降低,导致杀菌信号通路受阻。金黄色葡萄球菌通过脂磷壁酸(lipoteichoic acid,LTA)介导,靶向作用于TLR3激活的角质形成细胞,调控特定皮肤炎症通路。
皮肤微生态概述
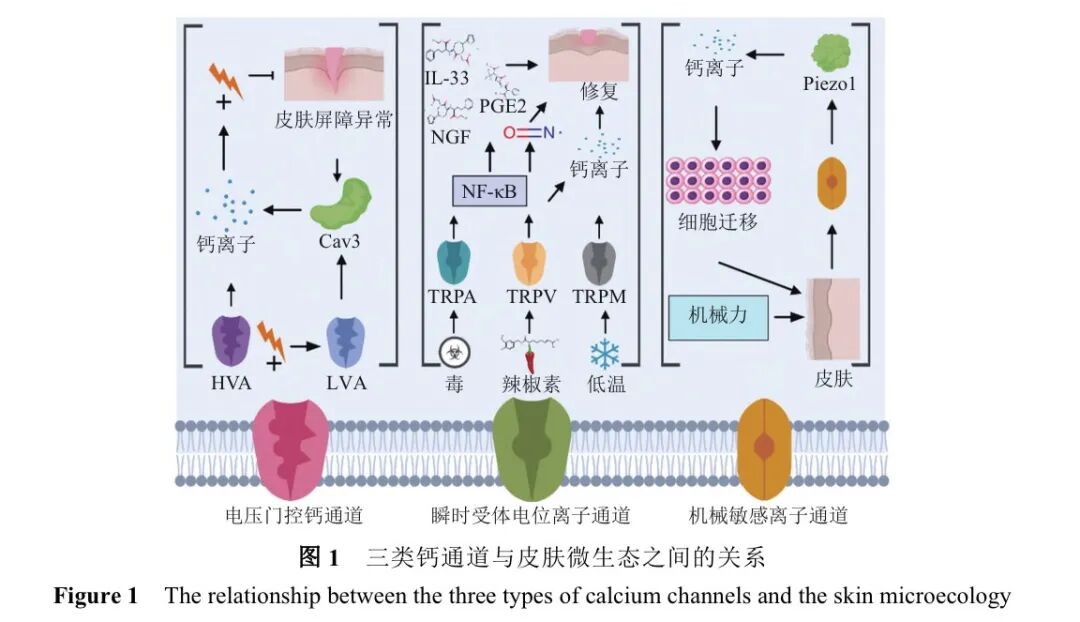
各种钙离子通道调节细胞内钙水平的动态波动,有助于维持皮肤微生态的平衡。钙离子通道主要包括VGCC、TRP和MS三类。这三类钙离子通道与皮肤微生态之间的关系如图1所示。
3.1、VGCC:
VGCC是一种跨膜蛋白,通常由膜电位 的去极化激活。根据其电压依赖性,VGCC通常可分为高压激活和低压激活,后者通常被称为T型通道,需要较小的膜电位去极化才能打开。T型通道的亚基为Cav3,其功能包括改变药理学特性、增加电流密度和调控钙离子的膜运输。此外,该亚基似乎还参与了去除钙离子的机制,以及对质膜中钙的回收或降解。重组Cav3通道表现出与天然T型电流相似的电生理活性,能够在较低电压下激活并更快失活。研究证实,Cav3在小鼠背根神经节 神经元的机械感应亚群中表达,缺乏Cav3会减少组胺、IL-4和胸腺基质淋巴细胞生成素(thymic stromal lymphopoietin,TSLP)诱导的瘙痒。敲除Cav3基因或使用T型钙通道阻滞剂mibefradil进行治疗,均减少了特应性皮炎模型小鼠的自发性及机械诱导的抓挠行为和皮肤炎症。而破坏IL-6信号传导受体亚基会阻断VGCC,皮肤间充质干细胞在炎症条件下会分泌高水平的IL-6,导致在间充质干细胞的迁移过程中出现连续黏附和脱黏附。或许VGCC通过依赖于炎性因子的机制,介导皮肤的超敏反应,以响应潜在的破坏性事件。
刺激磷脂酶C耦联受体并形成肌醇三磷酸后,表皮角质形成细胞上的VGCC减少了钙离子释放。当皮肤屏障受损时,钙离子进入细胞内后会产生正电位,从而延缓皮肤屏障的修复。VGCC能够调节外部电位对表皮中钙离子梯度的影响,从而影响皮肤屏障的稳态,因此在离子通道中占据特殊位置。VGCC能够将膜兴奋性转化为细胞质中钙离子浓度的变化,这是细胞对电信号反应的基础。皮肤溃疡、特应性皮炎和银屑病等疾病与此密切相关,深入研究有助于进一步了解皮肤微生态失衡的作用机制。
3.2、TRP:
TRP以该超家族的创始成员命名。这个超家族是果蝇光传导突变体表型的基础,直到1992年,果蝇的TRP通道功能才首次得到证实。在哺乳动物中,TRP家族包括TRPV、TRPC、TRPM、TRPA、TRPP、TRPML和TRPN。TRP超家族由一组不同的阳离子通道组成,4个TRP通道亚基可以组装形成功能性同源或异源四聚体。这些亚基向细胞内传递钙离子,调节角质形成细胞的生长和分化,从而影响皮肤的屏障功能。
TRPV对各种组织损伤信号敏感,其激活通常被认为与疼痛相关。表皮角质形成细胞上TRPV1的激活导致钙离子流入细胞,抑制细胞生长并诱导细胞凋亡。辣椒素作为TRPV1的激动剂,以特异性和剂量依赖的方式抑制细胞因子诱导的细胞分化、 细菌吞噬及促炎细胞因子的分泌。由于皮脂腺脂质有助于建立皮肤屏障,辣椒素对人皮脂腺细胞TRPV1的激活抑制了脂质的合成。一项研究证实,在未分化的人原代角质形成细胞中,抑制TRPV1和肌醇三磷酸受体会增加钙离子的储存、NF-κB的活性以及炎症介质的产生。TRPV1的信号传导可能会抑制表皮屏障功能并减少炎症介质的产生。TRPV2在结构上与TRPV1相似,但TRPV2中TRP结构域与S6之间的结构域间变构耦合较为松散。这种耦合的缺乏是由于缺少π螺旋,导致与TRPV1结构不同的失活构象。TRPV2抑制剂有效减弱了转化生长因子(transforming growth factor,TGF)-β信号传导、角质形成细胞释放的TGF-β和肌成纤维细胞α平滑肌肌动蛋白的产生而导致的角质形成细胞依赖性胶原细胞的凝胶方向性收缩。这或许有助于研究皮肤屏障受损和疤痕收缩的机制。
在其他TRPV通道中,TRPV3和TRPV4可能在皮肤功能的调节中发挥重要作用。激活TRPV3促使角质形成细胞释放一氧化氮(NO),NO是一种在包括伤口愈合在内的多种皮肤稳态机制中起重要作用的介质。同时,TRPV3还可以通过NF-κB通路促进炎症反应,增加人角质形成细胞中前列腺素E2、神经生长因子、TSLP和IL-33的产生。与TRPV3类似,TRPV4也参与表皮屏障的稳态调控。Kida等的研究表明,TRPV4的激活对于屏障的形成和恢复至关重要。通过激活TRPV4,钙离子能够从细胞外流入,从而促进细胞间连接的发育。这些变化导致体外细胞间屏障完整性的增强, 同时也减少了钙离子转化后细胞间渗透的增加。
TRPA家族只有一个成员TRPA1,存在于上皮细胞和毛细胞中。有害的外部刺激或低温会激活TRPA1。TRPA1能调节表皮细胞的生物学功能, 例如抑制TRPA1可以降低皮肤水肿和促炎细胞因子的水平。常见的拮抗剂包括布洛芬-酰基−葡萄糖醛酸苷和GDC-0334。TRPA1在皮肤组胺非依赖性瘙 痒中也发挥着主要作用,这一过程是许多慢性瘙痒疾病的基础。研究发现,TRPA1基因敲除时,特应性皮炎小鼠的瘙痒反应显著降低。通过组学分析发现特应性皮炎患者的TRPA1基因表达水平显著增加,推测这与IL-31在特应性皮炎瘙痒病变皮肤中的转录和表达水平上调有关。同时,TSLP也被发现与特应性皮炎相关。TSLP与其受体直接结合于神经元背根神经节,并通过磷脂酶C的下游信号传导,在TRPA1的协助下诱导瘙痒。这个过程由蛋白酶激活受体 2(protease-activated receptor 2,PAR2)介导,PAR2在角质形成细胞TSLP的产生中起重要作用。关于TRPA1还有其他发现,例如TRPA1激动剂可以促进皮肤屏障的恢复。考虑到特应性皮炎和微生物接触性皮炎均涉及表皮屏障功能障碍,这或许可以成为未来研究皮肤微生态平衡的一个关键靶点。
TRPM亚家族由8个可变成员组成,即TRPM1-8。除TRPM4和TRPM5外,TRPM通道是钙离子可渗透的阳离子通道,与TRPV1类似,更容易受到低温的影响。TRPM通道在体内钙离子平衡、感觉知觉和细胞信号传导等多种生理过程中发挥着关键作用。在调节TRPM通道活性的因素中,钙调蛋白作为重要的调节因子脱颖而出,其激活可以抑制结肠组织中降钙素基因相关肽的产生,并阻止化学诱导的刺激。一项研究表明,在炎症性肠病模型小鼠中,皮肤损伤激活了透明质酸的降解,导致结肠成纤维细胞功能的改变,进而通过产生抗菌肽来增强结肠中的炎症反应。上述研究表明,皮肤与其他屏障部位之间存在双向连接。
3.3、MS:
MS形成了一个不断扩展的离子门控通道家族,这些通道能够通过直接物理力或质膜的变形而被激活。钙可渗透性MS在肿瘤发生和发展中,可调节细胞迁移的起始、方向和持久性。维持皮肤微生态平衡涉及多个复杂的过程,包括炎症、组织增殖和屏障修复,而这些过程都伴随着机械力的持续作用。机械传导是皮肤接收来自内部和外部环境的物理信号并作出反应的机制,它将这些信号转化为细胞内的生化信号。这个复杂的过程依赖于一 种被称为机械换能器的特殊蛋白质。Zhu等的研究发现Piezo1是一个关键的MS,在上述过程中发挥着核心作用。冷冻电子显微镜揭示了Piezo1的三聚体螺旋桨状结构,其胞外结构域类似于三个远端叶片加上一个中央帽。这种三聚体结构表明,Piezo1可能将其外围区域用作力传感器,以调控钙离子的导电孔。进一步研究机械力调节Piezo1活性的机制发现,当膜处于闭合状态时,Piezo1会局部变形为圆顶形状。Piezo1从闭合状态转变为开放状态增强了其对机械刺激的响应能力,或许膜张力与圆顶下投影面积的变化会影响门控能量。
内皮中表达的Piezo1对剪切应力敏感,导致钙内流。当细胞内钙含量上升时,钙结合钙调蛋白后诱导CaMKⅡ磷酸化,调节免疫反应与慢性炎症。代表性的有来自内皮的抗菌肽在表皮葡萄球菌产生 的小分子引发的TLR2信号传导激活中增加,诱导β-防御素家族成员对皮肤中的大肠埃希菌(Escheri-chia coli)和金黄色葡萄球菌具有杀菌作用。除抗菌肽外,金黄色葡萄球菌还通过辅助基因调节因子来影响其在皮肤的定植,并介导上皮损伤和炎症,同时为其他共生微生物提供靶点,以抑制病原微生物的生长和毒性。Piezo1在修复皮肤损伤方面也具有一定的潜力。例如,低强度脉冲超声可以显著促进MC3T3-E1细胞迁移和增殖,但是敲除Piezo1限制了细胞的迁移和增殖。Jiang等的研究发现Piezo1-shRNA抑制Piezo1表达,从而抑制人骨肉瘤细胞侵袭。重建皮肤屏障的完整性至关重要,长期以来受损伤口的愈合、炎症状况和疼痛都困扰着所有生物。因此,通过Piezo1进行生物物理刺激可能是一种有效的治疗手段。
3.4、钙通道对皮肤微生态的调控机制:
钙离子通道对皮肤微生态的调控依赖于不同类型通道的协同作用。VGCC、TRP和MS通过信号交互共同维持皮肤微生态平衡。例如,VGCC中的Cav3激活后释放IL-4和TSLP等炎性因子,间接激活角质形成细胞的TRPV1通道,放大钙信号,而TRPV1通过钙依赖性磷酸酶抑制VGCC活性,防止炎症过度损伤共生菌。金黄色葡萄球菌的δ毒素可同时刺激这两种通道,扰乱抗菌肽分泌,破坏表皮葡萄球菌的竞争优势。机械拉伸激活Piezo1通道后触发钙内流,增强TRPV4活性并促进细胞连接蛋白表达,从而强化皮肤屏障功能,抑制痤疮丙酸杆菌的入侵。表皮葡萄球菌的短链脂肪酸通过抑制TRPA1活性,降低Piezo1对机械刺激的敏感性,减少共生菌的免疫损伤。在皮肤损伤修复过程中,三类通道分阶段协作:急性期VGCC驱动中性粒细胞清除病原菌;增殖期TRPV3感知局部温度升高,促进角质细胞迁移并招募调节性T细胞;重塑期Piezo1通过基质刚度变化激活胶原沉积,同时上调抗菌肽,抑制耐药菌定植。另一方面,微生物群也反向调控通道活性,例如表皮葡萄球菌分泌的PSMγ蛋白抑制TRPV1过度激活,而铜绿假单胞菌劫持VGCC信号促进自身存活。微生物群与调控通道相互作用,共同维系皮肤微生态平衡。
总结与展望
本文阐述了一些常见的钙离子通道参与皮肤屏障修复的机制,以及在控制细胞生长和分化中的作用。同时还指出钙离子参与免疫或炎症反应,并影响某些皮肤微生物的定植。然而,对于皮肤微生态失衡导致的一些疾病,仍缺乏大量的临床针对性研究。借助单细胞空间转录组技术和细胞索引测序的联合应用,未来可精确解析表皮基底层、棘层和颗粒层细胞中钙离子通道蛋白的动态表达谱,这将为阐明皮肤免疫微环境的细胞异质性提供全新视角。微生物荧光原位杂交与高分辨率钙成像技术的整合策略,使得在活体水平同步监测金黄色葡萄球菌生物膜形成和宿主细胞钙振荡成为可能,此类实时动态数据将显著提升对微生物−宿主信号传导耦联机制的理解深度。
参考文献:
1.董汭,阮海华,武陶,张宏扬.钙离子通道影响皮肤微生态机制研究进展[J].中国微生态学杂志,2026,1(38):113-119.
2.Borrego-Ruiz A, Borrego JJ.Microbial dysbiosis in the skin microbiome and its psychological consequences[J]. Microorganisms, 2024, 12(9): 1908.
3.Skowron K, Bauza-Kaszewska J, Kraszewska Z, et al. Human skin microbiome: impact of intrinsic and extrinsic factors on skin microbiota[J]. Microorganisms, 2021, 9(3): 543.
4.其他文献略。
来源:海龙话皮微信公众平台
医脉通是专业的在线医生平台,“感知世界医学脉搏,助力中国临床决策”是平台的使命。医脉通旗下拥有「临床指南」「用药参考」「医学文献王」「医知源」「e研通」「e脉播」等系列产品,全面满足医学工作者临床决策、获取新知及提升科研效率等方面的需求。
 我要投稿
我要投稿














{kind=link}