
近日,Vera Adema及Simona Colla团队在《Blood Cancer Discovery》发表综述,医脉通特此系统梳理了稳态下红系造血的调控网络,深入解析了MDS贫血的分子机制,并评估了当前针对低危MDS贫血治疗策略的作用机制与临床证据,为该领域的个体化治疗提供了重要参考。
稳态红系造血:多层次调控下的复杂分化过程
成人红系造血起始于造血干细胞向巨核-红系祖细胞的分化,随后依次分化为早期红系祖细胞和终末红系分化阶段。终末红系分化不依赖促
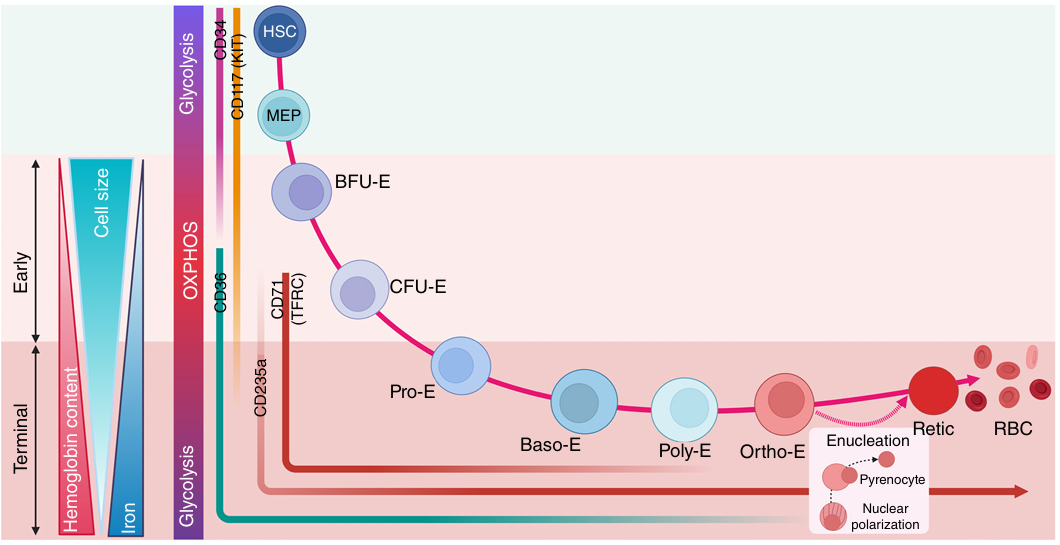
图1 红系造血示意图
红系分化可通过流式细胞术结合CD71、CD105、CD235a等表面标志物进行阶段特异性分析。该过程受到外源性因子与内源性转录因子的严密调控。
外源性调控:SCF、EPO与红系岛微环境
干细胞因子通过结合c-Kit受体,激活PI3K/AKT及MAPK信号通路,促进早期红系祖细胞的存活与增殖。EPO作为红系造核心激素,由肾脏分泌,结合EPO受体后激活JAK2/STAT5等信号通路,驱动红系细胞存活、增殖与成熟。EPO信号受SHP-1及SOCS家族负反馈调控,防止红细胞过度生成。
红系岛是骨髓中特化的造血微环境,由中央巨噬细胞与不同阶段红系细胞构成。巨噬细胞提供细胞因子、铁离子并吞噬脱出的细胞核,并通过VCAM-1、ICAM-4等黏附分子维持结构完整,支持红系分化(图2)。
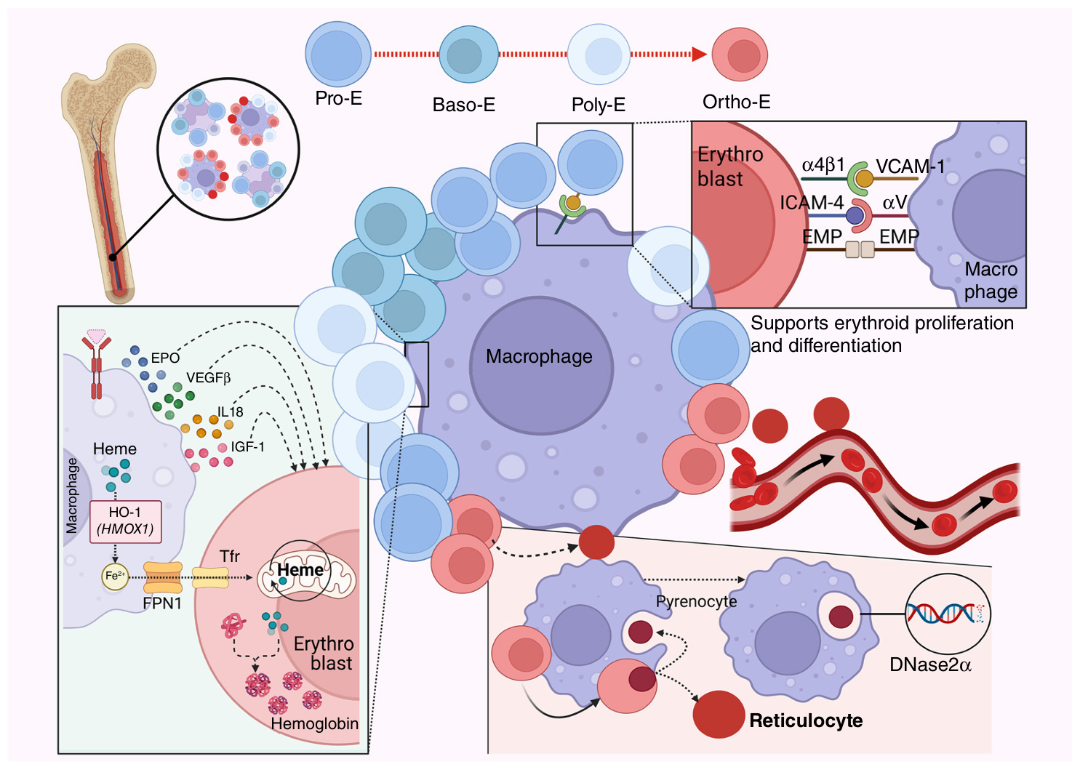
图2 红系岛结构示意图
内源性转录因子:GATA1、KLF1与TAL1的核心作用
GATA1是红系与巨核系发育的主转录因子,在MEP阶段通过“GATA转换”启动红系定向分化。GATA1调控珠蛋白、血红素合成、自噬及红细胞膜相关基因表达。KLF1调控红系定向、血红蛋白转换及终末分化,并激活BCL11A以抑制γ-珠蛋白表达。TAL1与GATA1、KLF1协同调控终末红系分化相关基因。
线粒体与代谢:能量与血红素合成的整合节点
红系造血过程中,细胞从早期依赖氧化磷酸化转变为终末阶段依赖糖酵解。GLUT1表达上调以维持无氧糖酵解。血红素合成始于线粒体,涉及ALAS2等关键酶,终产物与珠蛋白结合形成功能性血红蛋白。铁通过TFR1内化、还原及转运进入线粒体,由FECH插入原卟啉IX生成血红素。EIF2AK1通路在血红素充足或缺乏时调控珠蛋白翻译(图3)。
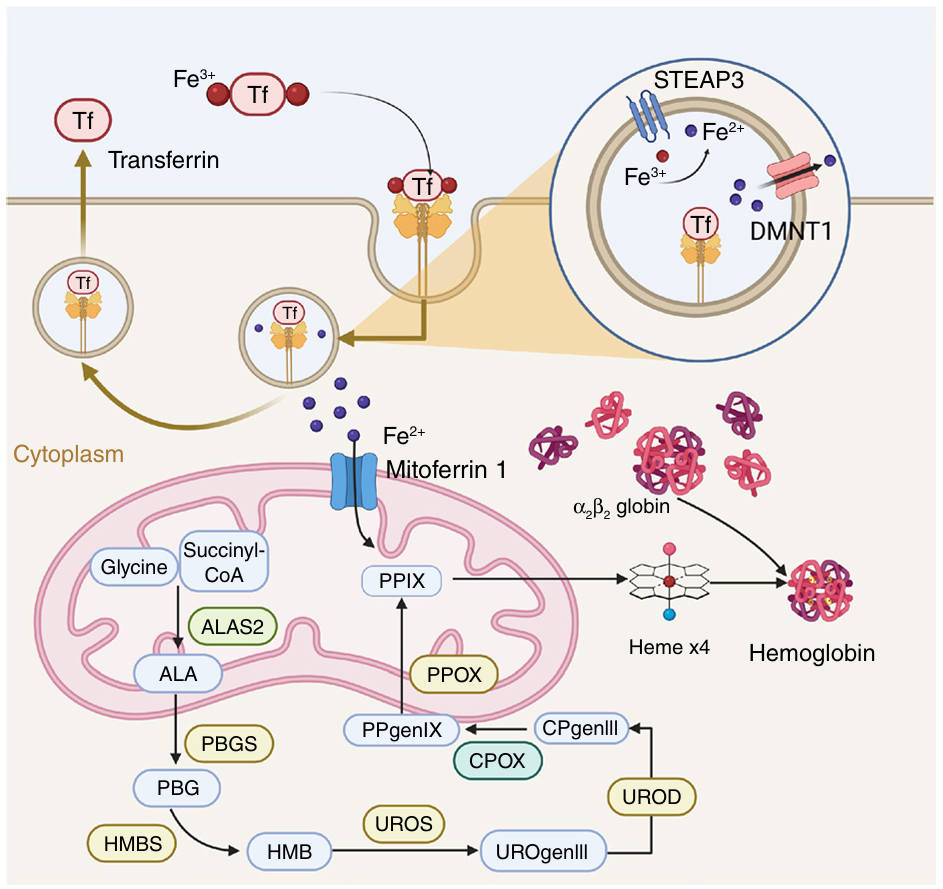
图3 血红素合成与血红蛋白生成示意图
MDS贫血机制:无效红系造血的分子基础
MDS-del(5q)中,RPS14单倍剂量不足激活p53依赖性红系凋亡,CSNK1A1单倍剂量不足增强
高危MDS贫血则源于造血干细胞向髓系分化偏离,而非单一突变导致的红系阻滞,治疗重点在于延缓疾病进展而非直接纠正贫血。
低危MDS贫血治疗策略:从ESAs到新型靶向药物
促红细胞生成素类药物(ESAs)为一线治疗,适用于内源性EPO水平较低、
Imetelstat为
来那度胺在MDS-del(5q)中通过诱导CK1α降解实现合成致死。MDS-004研究显示,10mg来那度胺组61%患者脱离输血,中位反应持续时间未达到。在非del(5q)患者中亦有一定疗效,但需警惕粒细胞减少与血小板减少。
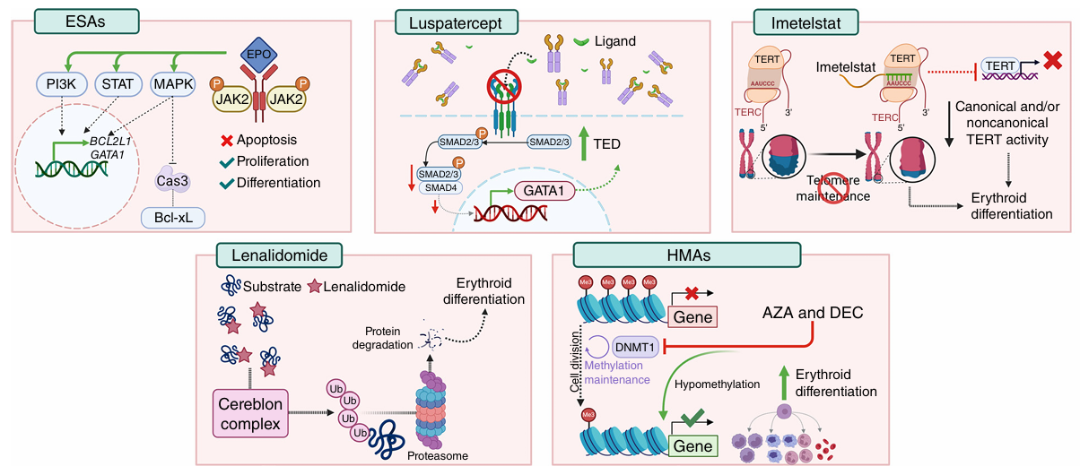
图4 低危MDS贫血治疗药物的作用机制示意图
未来方向:从机制到精准干预的探索
总结:理解无效红系造血,推动MDS贫血个体化治疗
参考文献:
Adema V, et al. Understanding the Molecular Mechanisms Underlying Anemia in Myelodysplastic Syndromes: From Erythropoiesis to New Therapeutic Approaches. Blood Cancer Discov. 2026 May 6:OF1-OF12.
编辑:Felicia
审校:Janet
排版:Wyn
执行:Baa
 我要投稿
我要投稿













{kind=link}